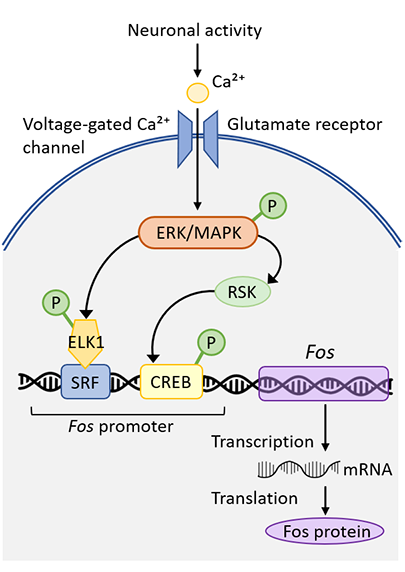
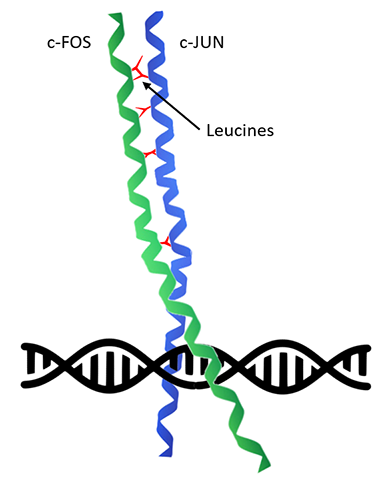
Indeed, c-FOS “bridges the gap” by mediating the conversion of extracellular signals into changes in gene expression. For instance, as depicted in Figure 1, extracellular triggers like the excitatory neurotransmitter glutamate or openers of voltage-gated calcium channels can generate an intracellular cascade that, via activation of kinases, results in the phosphorylation and binding of phospho-ELK1 and phospo-CREB to fos gene promoter regions that drive up-regulation of c-FOS protein levels. Due to the rapid increase in levels and early response of this proto-oncogene, c-fos has been dubbed an ‘immediate early gene’ as an early responder to various cellular perturbations including electrical stimulation, exposure to serum, growth factors, and neurotransmitters.